检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
 下载Firefox
下载Firefox
检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
下载Firefox
/ 6165金沙总站官方入口 / 学术科研
2020年2月4日,《自然·通讯》(Nature Communications)在线发表了北大6165cc金沙总站检测中心、北大麦戈文脑科学研究所、北大-清华生命科学联合中心唐世明课题组的研究论文:Spatiotemporal functional organization of excitatory synaptic inputs onto macaque V1 neurons(猕猴初级视觉皮层(V1)神经元树突上兴奋性输入的时空功能组织)。该研究借助于新型谷氨酸探针iGluSnFR,实现了对清醒猕猴大脑皮层神经元的双光子树突成像,并获得了单个V1神经元树突上兴奋性输入的精细时空功能图谱。
神经元作为神经系统最基本的计算单元,从它们的树突上接收信号的输入。单个神经元可以通过树突棘接收数以千计甚至数十万的突触前输入,加上树突的复杂几何构型以及其多样化的生理特性,使神经元具有对其输入信号进行复杂计算的能力 (Stuart and Spruston, 2015)。从上世纪末本世纪初开始,树突整合机制的相关研究已经逐步开展,这些研究采用离体的膜片钳记录手段和胞内记录,对单个神经元树突上突触输入的整合方式及其生理基础进行了有效且深入的探索 (Cash and Yuste, 1999 , Polsky et al., 2004)。树突上输入信号的线性与非线性整合,以及输入时间序列在树突计算中承载的重要作用等结果的发现,不断加深和拓展了我们对神经元树突功能的认知 (Larkum et al., 1999 , Branco et al., 2010)。然而,离体的单细胞或是脑片实验终究无法模拟真实的神经网络所承载的生理功能,如各类体觉感知以及高级认知。为了揭示树突整合机制所承载的功能效应,研究者们同样借助于传统却高保真的电生理技术手段进行了一系列的在体研究,进一步解释了树突活动对皮层功能的影响,例如视觉系统中的朝向和运动方向选择性 (Smith et al., 2013 , Wilson et al., 2018)。即便如此,传统的电生理手段仍极大地受限于其采样量和空间分辨率的大小,致使其在树突相关研究中的实用性远远无法满足该领域的诉求。
在本世纪第二个十年的开端,Konnerth课题组的一系列研究开启了树突成像的研究热潮 (Jia et al., 2010 , Chen et al., 2011 , Chen et al., 2013)。借助具有高空间分辨率的双光子成像手段,他们实现了对完整大脑中树突输入长时程的、具有单突触精度的功能测量。自此,一系列在体的树突成像研究不断地丰富了我们对树突输入的功能组织、计算机制及其生理意义的认知 (Takahashi et al., 2012 , Wilson et al., 2016 , Iacaruso et al., 2017 , Scholl et al., 2017 , Varga et al., 2011),其中令人兴奋的发现包括但不局限于:树突上的突触输入在功能上有空间聚类的特点、单神经元水平上输入与输出信号的关联与差异、视觉认知中朝向与运动方向选择性的树突机制等等。尽管如此,相关的研究都是借助钙信号探针在麻醉状态下的低等哺乳类动物中进行,该应用场景仍然不够理想,概括说来其不足共有三点:其一,钙信号树突成像中反向传播的全局动作电位 (back-propagating spikes, BAPs) 是极大的且无可避免的噪声源,即便经过有效的后期处理也会在很大程度上影响输入信号的纯净度;其二,考虑到麻醉对于神经元自发放电率的显著影响以及对树突膜特性的抑制效应 (Stuart and Spruston, 2015),当下的研究无法很好地模拟大脑真实的生理活性;其三,低等哺乳类动物在色觉、视觉敏感度和视觉认知方面的处理能力较为低下 (Roelfsema and Treue, 2014),很难帮助我们理解精细而复杂的高级视觉认知机制。
基于上述研究现状,本研究借助于新型谷氨酸探针iGluSnFR实现了高质量的树突成像,使得我们可以对清醒猕猴大脑皮层神经元树突上的突触输入进行精确的功能测量,并绘制了单个V1神经元树突上兴奋性输入的精细时空功能图谱(图1a)。我们发现,在单个V1神经元的底树突上,朝向和颜色选择性输入具有功能上的整合和竞争,这为V1浅层神经元中朝向与颜色信息的联合编码机制提供了树突水平上的直接证据(图1b)。值得一提的是,单个神经元接收到的朝向选择性输入的比例越高,它们的一致性也就越强,这可以用于解释V1神经元朝向选择性强弱的由来 (Wilson et al., 2016 , Takahashi et al., 2012)。
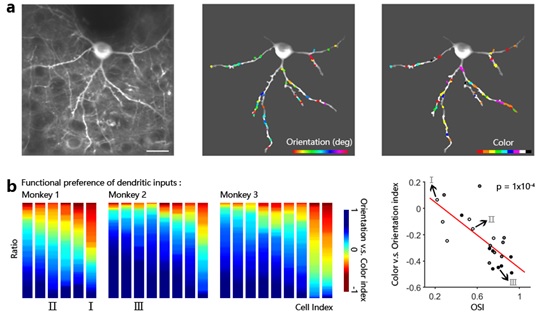
图1. 猕猴V1神经元树突上的兴奋性输入。
a,单个V1神经元树突上兴奋性输入的空间分布;b,朝向和颜色选择性输入在单个V1神经元底树突上的整合与竞争。
对单个视觉特征而言,树突上的输入倾向于形成空间聚类,然而在多维特征空间中情况却并非如此。树突上的每个输入都有着特定的特征偏好组合(包括朝向、空间频率和感受野),这些组合在多维特征空间中散乱分布。同一树突分支上的一对输入可能拥有相同的朝向偏好性和不同的感受野(图2a);相反,另一对输入可能同时拥有不同的朝向偏好性和不同的感受野(图2b),或者不同的朝向偏好性和相同的感受野。树突的特征整合能力依赖于其输入组合的丰富性,上述这一特性正有助于最大化其潜在的特征组合池(图2)。正因如此,它为V1浅层神经元提供了树突水平上多维特征整合的潜在计算基础。

图2. 树突输入在多维特征空间中功能分散。
a,一段树突枝干上的两个输入,有相同的朝向偏好和不同的感受野;b,另一对输入,有不同的朝向偏好和不同的感受野。
我们还发现,相比于底树突,顶树突上的输入具有更大的感受野和更长的反应延迟,进一步印证了顶树突接收反馈信号的输入,而底树突则接收视觉信息流中的前馈输入 (Marques et al., 2018 , Zhang et al., 2018)。但是,在实验中给出反馈和前馈输入的直接证据仍然是一个挑战,而我们的结果仅仅是为信号的来源提供了间接的证据。我们接下来的研究正是为了改变这一现状,旨在通过结合新技术(例如光遗传学或电生理学)和特定的行为任务(例如自上而下调节的选择性注意任务)来获得直接的证据。我们的功能性测量与实验结果,结合首次在非人类灵长类动物中成功应用新型谷氨酸探针,为进一步理解视觉信息处理的树突机制和计算原理提供了桥梁。
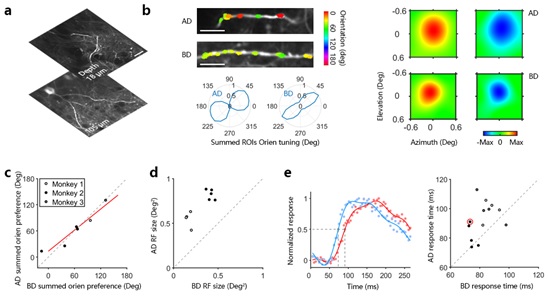
图3. 突触输入在顶树突和底树突上不同的功能组织。
a,示例神经元的顶树突与底树突;b,顶树突与底树突上输入的功能测量;c,顶树突与底树突上的输入和具有相似的朝向偏好;d,顶树突上的输入具有更大的感受野;e,顶树突上的输入有更长的反应延迟。
6165cc金沙总站检测中心、北大麦戈文脑科学研究所、北大-清华生命科学联合中心唐世明研究员为该论文的通讯作者。6165cc金沙总站检测中心2015级博士研究生居年盛为该论文的第一作者。唐世明课题组技术员李扬、刘芳、姜鸿飞,纽约州立大学Stephen L. Macknik和Susana Martinez-Conde教授也参与该研究的部分工作。该研究工作得到了国家自然科学基金、国家杰出青年科学基金、国家重点基础研究发展规划项目和国家985重点建设项目的资助。
原文链接:https://www.nature.com/articles/s41467-020-14501-y
地址:北京市海淀区颐和园路5号
金光生命科学大楼
电话:010-62757794

北大生科官方微信

生声不息公众号