检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
 下载Firefox
下载Firefox
检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
下载Firefox
/ 6165金沙总站官方入口 / 学术科研
2023年11月21日,6165cc金沙总站检测中心陆剑课题组与合作者在《Biological Reviews》上发表了题为“Convergent and divergent evolution of microRNA-mediated regulation in metazoans”的综述论文。
microRNAs(miRNAs)是一种小的非编码RNA,长度通常在20到24个核苷酸之间,在植物和动物中表达,通过破坏靶mRNA的稳定或抑制其翻译来调节靶基因的表达。microRNA和靶位点怎样起源,怎样演化?陆剑与合作者先前围绕上述问题取得了一系列原创性成果,提出新microRNA快速产生与死亡的观点(Lu et al., 2008, Nature Genetics),阐明适应性进化如何驱动新microRNA产生功能(Lu et al., 2008, MBE),揭示了microRNA调控对人类进化的影响(Lu et al., 2012, Genome Research)。近年来提出miRNA起源的“功能共适应”模型,阐述自然选择如何驱使同一簇中的microRNA进化出相近的功能(Wang et al., 2016, MBE); 发现microRNA复制后在表达和功能上产生广泛的分化,提出microRNA复制促进新靶位点功能建立这一崭新模型(Luo et al., 2018, RNA); 发现真核生物5’ UTR中的上游读码框(uORF)和3’UTR中的microRNA靶位点在数量上显著正相关,呈共进化趋势,提出uORF和microRNA能够通过“失效保护”(fail-safe)机制,共同调控靶基因mRNA的稳定性或翻译效率(Zhang et al., 2021, Science Bulletin)。在此综述中,作者总结该领域相关进展,探讨了miRNA调控途径的起源与演化模式,总结并概述了在两侧对称动物中自然选择对miRNA演化的驱动,阐明了两侧对称动物中miRNA与其潜在靶标的共同演化规律,旨在增进我们对两侧对称动物miRNA的功能和演化动态的理解。
虽然在植物和动物中都发现了miRNA,但它们在生物发生过程和功能机制方面存在显著差异。一种假说认为,植物和动物中的miRNA途径独立于RNAi途径进化而来,这意味着现代miRNA是通过趋同进化而出现的。然而,最近来自非两侧对称后生动物、原生生物和植物的证据提出了另一种可能性,即miRNA途径可能起源于一个共同祖先(图1)。动物和植物之间miRNA途径和miRNA库的差异主要源于谱系特异性miRNA途径革新、miRNA途径缺失、miRNA获取以及与真核生物最后共同祖先分化后miRNA的缺失。
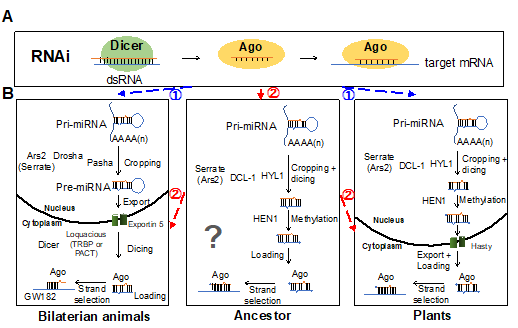
图1. 动植物miRNA通路的进化起源
miRNA通路中关键组分的保守模式有助于深入了解miRNA通路的进化史(图2)。AGO和Dicer在各种真核生物中的存在支持了真核生物的最近共同祖先拥有RNAi通路的观点。ARS2在动物和植物中均存在(在植物中称为Serrate)。尽管Drosha和Pasha在植物中不存在,但它们起源于后生动物共同祖先之前。这些数据表明,原始动物祖先具有类似于植物和刺胞动物祖先类型的miRNA通路。值得注意的是,在两侧对称动物中,miRNA通路发生了革新,动物Drosha可能是通过复制Dicer进化而来的。因此,在两侧对称动物的原始祖先中,Drosha/ARS2/Pasha复合物在处理miRNA前体方面的功能可能类似于DCL/Serrate/HYL1。此外,Loqs基因家族起源于后生动物,并与Dicer合作在细胞质中处理pre-miRNA。因此,对miRNA生物发生机制的进化分析支持了miRNA途径起源于“共同祖先”的假说。
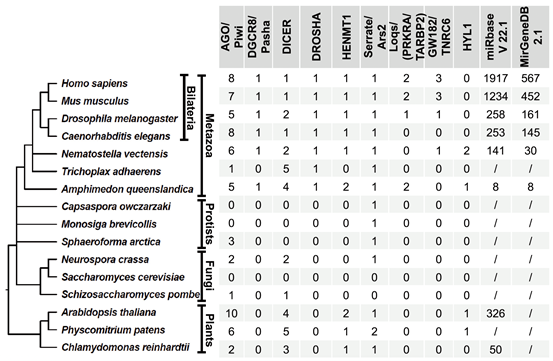
图2. 真核生物中miRNA通路的同源蛋白和已注释的miRNA数目
miRNA革新与形态复杂性之间可能存在联系。图3显示了基于MirGeneDB的系统发育树中miRNA家族和基因的得失。值得注意的是,在脊椎动物进化到现代人类的过程中,有四个进化分支经历了大量miRNA家族的增加。首先,在无脊椎动物和脊椎动物分化之前,获得了32个家族(包含39个miRNA基因)。其次,在脊椎动物的底部,增加了45个新家族(176个miRNA基因)。第三,在真兽亚纲的哺乳动物基础上,获得了91个家族(144个miRNA基因)。最后,在人类与小鼠分离后,获得了111个家族(179个miRNA基因)。总的来说,这些观察结果强调了在两侧对称动物的进化史中,miRNA基因的持续获取,以及在特定动物谱系中miRNA革新的显著提高。
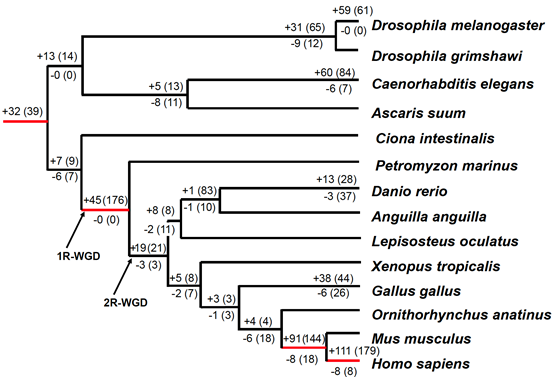
图3. miRNA在两侧对称动物进化过程中的得失
早期的研究主要使用低通量测序方法,集中研究进化上保守的miRNA。这些miRNA被认为在功能上很重要,并且在各个物种中常常表现出高水平的表达。因此,不同物种间miRNA序列的保守性被视为miRNA鉴定的重要标准。然而,随着二代测序技术的进步,发现了大量进化上不保守或谱系特异性的miRNA。非保守的miRNA可以通过转录基因组区域内的新形成来产生(图4A)。然而,新形成的miRNA可能会无意中干扰转录组或翻译过程,产生有害影响,导致它们最终被自然选择从种群中清除。因此,在强大的选择压力下,许多新的miRNA在进化中是短暂存在的,并且对特定的谱系具有特异性。复制是促进新miRNA产生的另一种机制。miRNA复制事件可以局部发生在基因组的不同区域,也可以通过全基因组复制事件发生(图4B)。此外,刺胞动物表现出miRNA前体由其靶位点的反向复制产生的机制,类似于植物中观察到的过程(图4C)。
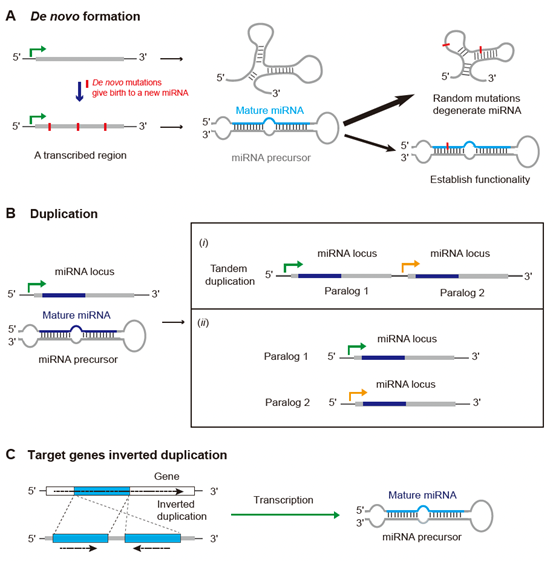
图4. miRNA诞生和死亡的分子机制
在两侧对称动物中,miRNA介导的调控通过多种机制呈现趋同进化。miRNA的聚类为新生miRNA提供了生存优势,并使其获得与先前存在的miRNA相似的功能。聚类中的miRNA倾向于在空间或时间上共表达,这使得新的miRNA逐渐与同一簇中先前存在的miRNA共享靶基因(图5A)。另外,同一簇中多个新形成的miRNA可能相互作用以调节重叠的靶基因集(图5A)。群体遗传学研究表明,正选择推动了果蝇和灵长类动物miRNA聚类的建立和进化。miRNA中模仿种子序列的趋同分子进化并不罕见(图5B),可能在基因调控中发挥关键作用。此外,不同miRNA介导的基因调控可导致相似的功能结果,这暗示了趋同的分子进化。举例来说,在两侧对称动物的发育过程中,参与母体mRNA降解的miRNA表现出不同的组成和序列(图5D)。
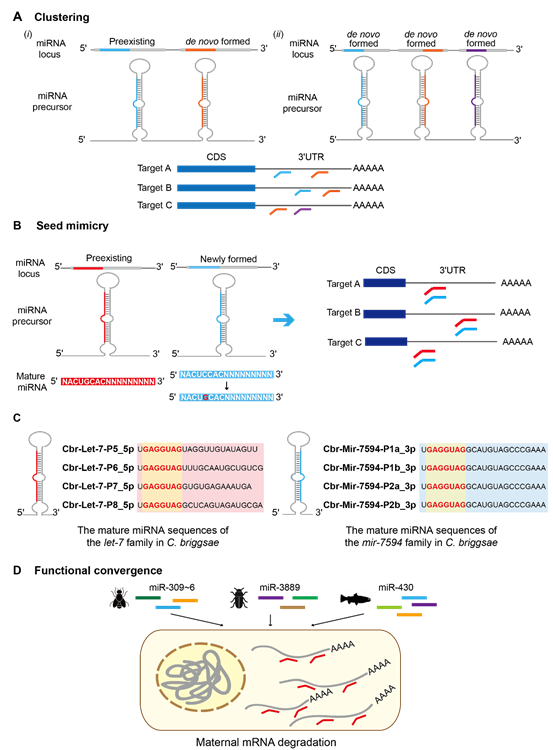
图5. miRNA介导的基因调控趋同进化
保守的miRNA通常具有典型的功能性和广泛的表达。然而,它们也可以通过种子或成熟miRNA序列的变化、时空表达模式的改变以及种子移动或手臂切换来促进物种之间的功能多样化(图6)。此外,即使是同源的miRNA,在远亲物种中其靶位点可能也存在差异。
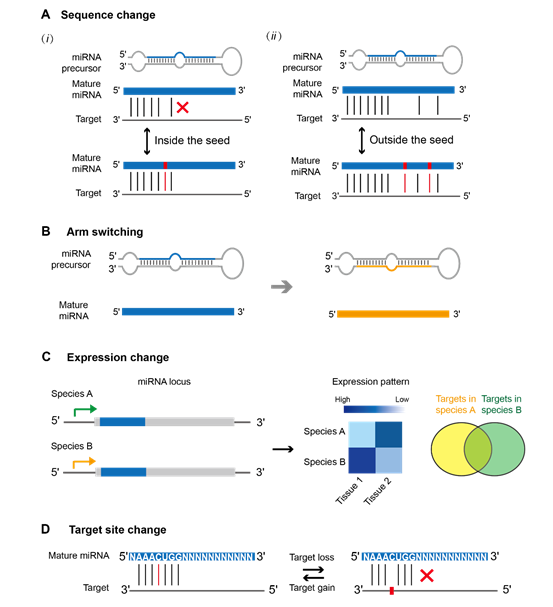
图6. miRNA介导的基因调控在物种间的分歧进化
miRNA复制后,种子序列、成熟序列之外的区域以及调控miRNA表达模式和加工效率的调控元件都会发生变化(图7A)。这些机制导致产生多种具有扩展功能的miRNA类似物。复制的miRNA可能展现出时间或空间上的表达差异,从而影响调控不同的靶基因集(图7B)。相较于单拷贝的miRNA,复制的miRNA具有更大的灵活性,可以在种子区域之外进行序列变化,以优化与新出现的靶位点的匹配(图7C)。因此,miRNA的复制有助于招募新的靶位点。
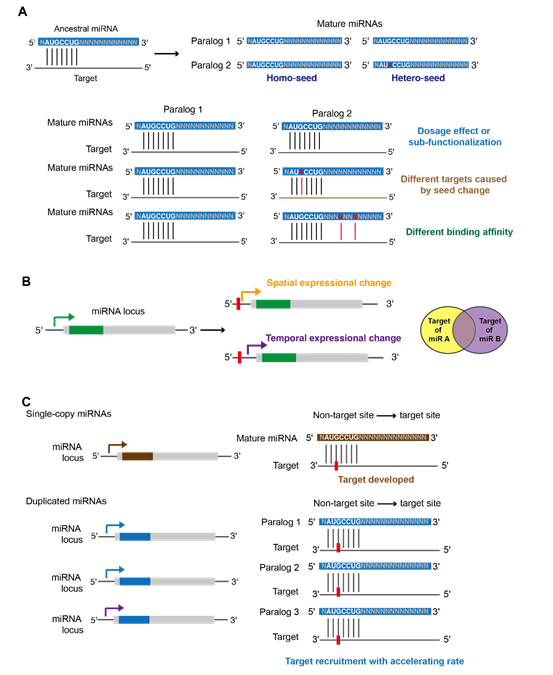
图7. 同源miRNA在复制后的功能多样化
湖南大学生物学院生物信息学中心王奕蓉副教授是文章的第一作者;王奕蓉及陆剑教授是文章的共同通讯作者; 6165cc金沙总站检测中心博士后唐小鹿对该论文做出重要贡献。该项研究得到了国家自然科学基金委、湖南省自然科学基金、中央高校基本科研专项资金、北京大学生科启东创新基金,以及蛋白质与植物基因研究国家重点实验室开放课题的资助。
文章链接: https://doi.org/10.1111/brv.13033
参考文献
1. Luo J#, Wang Y#, Yuan J#, Zhao Z, Lu J* (2018) MicroRNA duplication accelerates the recruitment of new targets during vertebrate evolution. RNA 24(6): 787–802.
2. Wang Y, Luo J, Zhang H, Lu J* (2016) MicroRNAs in the same clusters evolve to coordinately regulate functionally related genes. Molecular Biology and Evolution 33(9): 2232–2247.
地址:北京市海淀区颐和园路5号
金光生命科学大楼
电话:010-62757794

北大生科官方微信

生声不息公众号